The Upper (or Late) Triassic is when the action really starts to pick up. The Carnian Stage, the bottom of the Upper Triassic, begins where the ammonite species
Daxatina canadensis first appears; the top of the Carnian is at the base of the ammonite biozone of
Klamathies macrolobatus (or
Stikinoceras kerri, depending on whom you talk to). It’s during the Carnian that we meet the earliest sure-thing dinosaur,
Eoraptor, who appeared around 230 mya. The oldest well-documented dinosaurian assemblage, in the Ischigualasto Formation in Argentina, probably dates to the Late Carnian. It’s during this epoch that archosaurs truly begin to dominate the landscape, evolving into groups such as the phytosaurs, rhynchosaurs, aetosaurs, and rauisuchians. The first pterosaur,
Carniadactylus, appears; though pterosaurs started small, they would grow and diversify and dominate the Mesozoic skies. The therapsids, under the paw of the archosaurs, began to decrease in size and diversity; they would remain small and insignificant until the extinction of the dinosaurs (they would get their sweet revenge in the Cenozoic Era).
 |
| three common Shastasaurus species |
We look first at developments in the ocean, particularly with the ichthyosaurs. The pinnacle of ichthyosaur dominance and diversity was in the Upper Triassic, and in the Carnian we see several large species emerge.
Shonisaurus could reach up to fifty feet in length and had a long snout and flippers that were longer and narrower than those in most ichthyosaurs. Its body was more slender than most ichthyosaurs. Even larger than
Shonisaurus was
Shastasaurus and its kin. The shastasaurs were highly specialized and extremely slender in profile (the largest specimens had a rib-cage slightly less than six and a half feet deep despite a distance of over twenty-three feet between their flippers!).
Shastasaurus had a short, toothless snout, and it's been proposed that it was a ram-feeder, scouring the ocean floor and devouring everything kicked up in its path. The only species with several preserved skulls had a skull that measured only about eight percent of the body length. Interestingly, even babies lacked teeth (unlike
Shonisaurus juveniles, who had front teeth but lost them in adulthood).
Shastasaurus had a highly-compressed snout due to a unique arrangement of its skull bones. Numerous species have been identified, making this a predominant feature of the Upper Triassic ecosystem. A third species of ichthyosaur,
Cymbospondylus, was the 'apex predator' of the Upper Triassic. This ichthyosaur was somewhat eel-like in appearance and could reach up to thirty feet long. It lacked a dorsal fin and fluked tail but had an elongated snout like other ichthyosaurs. The head was about three feet long with large jaws and teeth designed for catching and holding fish and cephalopods (such as ammonites). Its long tail made up half its body length;
Cymbospondylus may have used its tail for locomotion in the same manner as modern sea snakes. It probably swam by wriggling its body side-to-side; the paddle-like fins would've served as stabilizers or brakes. It would've been quick and agile in the water, able to easily hunt fast-moving prey and tear into schools of fish. Adults probably spent much of their time hunting in the deep waters offshore, venturing into shallower water to breed or catch seasonal prey.
 |
| a pair of hunting Cymbospondylus |
 |
| Metoposaurus, a super-sized amphibian |
On land, supersized amphibians continued to diversify.
Metoposaurus was mostly aquatic with small, weak limbs, sharp teeth, and a large, flat head. This highly-flattened amphibian fed mainly on fish that it captured with its wide jaws lined with needle-sharp teeth. It grew up to ten feet long and clocked in around a thousand pounds. They may have used their limbs as flippers and swam in the same manner as the later plesiosaurs. The flat head and arm bones, wide hands, and large tail of
Metaposaurus diagnosticus implies that the creature swam in ephemeral lakes during the wet season and used its head and forearms to burrow underground when the dry season began.
Metoposaurus may have been preyed upon by the carnivorous archosaurian phytosaurs that lived in the same locales.
 |
| the crocodile-esque amphibian Koskinodon |
Another amphibian,
Koskinodon, could have reached up to ten feet in length. It was designed as an ambush hunter, snapping up anything small enough to fit in its huge jaws. Its teeth were designed to capture prey and were optimized for piercing flesh and keeping it from escaping. It had a wide, wedge-shaped, powerful tail to assist with swimming and defense. In some ways it resembled (and lived) like modern crocodiles, though it didn't reach near the same lengths and was rather short and stocky. Its stumpy legs show it had a sprawling stance, and it likely spent most of its time motionless, waiting for prey (it wouldn't need long legs to propel it long distances if it were an ambusher). It likely lay at the bottom of a shallow swamp, waiting for fish, crustaceans, smaller amphibians, or even young archosaurs to wander by. When it spotted prey, it would use its massive jaws to engulf them. An aquatic lifestyle is further suggested by the sensory sulci, useful for detecting changes in water pressure made by the swimming motions of nearby organisms. Mass graves of
Koskinodon have been discovered, probably the result of these animals being forced to herd together in shrinking swamps during a drought and perishing when the water dried up (quite like its older amphibian relative,
Mastodonsaurus, addressed earlier).
 |
| a family of Placerias at the water's edge |
Therapsids began losing steam against the rising archosaurs, but there were bright spots nonetheless.
Exaeretodon of the Ladinian continued into the Carnian, and two larger-than-life dicynodonts emerged:
Ischigualastia and
Placerias.
Ischigualastia was a large herbivorous dicynodont that could weigh up to two tons and which competed for space with
Hyperodapedon and
Exaeretodon. Though it was common in the early Carnian, it was whittled away to extinction, perhaps by being out-competed by other dicynodonts and being preyed upon by archosaurs and early dinosaurs.
Ischigualastia’s niche was taken over by another large (but somewhat less bulky) dicynodont
Placerias.
Placerias belonged to the last off-branching group of dicynodonts before dicynodonts became extinct at the end of the Triassic.
Placerias was one of the largest herbivores in the Upper Triassic, reaching up to eleven feet in length and weighing up to a ton. It had a powerful neck, strong limbs, and a barrel-shaped body. Most scientists believe it lived much like a modern hippopotamus, spending much of its time during the wet season wallowing in the water and chewing at bank-side vegetation. Such a lifestyle would’ve offered it protection against terrestrial predators like the emergent rauisuchians (see
Postosuchus below), and it could’ve put up a decent fight against phytosaurs and crocodylomorphs with its two short tusks and powerful beak.
 |
| the shrew-like Adelobasileus |
Although some paleontologists speculate that
Cynognathus may have been the first mammal (or at least ancestor to all mammals), not all agree. Others place the Carnian
Adelobasileus in this place of honor, arguing that the shrew-like
Adelobasileus was the common ancestor of all modern mammals or a close relative to that ancestor. This mammal-like synapsid (a group of animals that includes mammals and every animal more closely related to mammals than to reptiles and birds) is known only from a partial skull – but this partial skull is quite the find. Distinct cranial features, particularly the housing of the cochlea (part of the inner ear), suggests that this creature was a transitional form between therapsid cynodonts and Triassic mammals. Though it is not a ‘true mammal’ based upon trait-based taxonomy, many see it is as an early mammal – and thus one of humankind’s ancestors.
 |
| a pair of aquatic Tanystropheus |
Archosaurimorphs and archosauriformes (not-quite-archosaurs) continued to evolve despite their true archosaur cousins taking center stage. The aquatic tanystrophids continued to evolve, and
Tanystropheus (briefly mentioned during our foray into the Olenekian) emerged (though recent finds may place its emergence in the Ladinian of the Early Triassic).
Tanystropheus was about twenty feet long; its slender neck was larger than its body and tail combined. Because the neck was stiff, and because it's been found in semi-aquatic fossil beds, it's argued that it lived an aquatic or semi-aquatic lifestyle. Paleontologists differ (and, at times, do so strongly) on whether it was fully aquatic or semi-aquatic. It was piscivorous (fish-eating): its teeth were long, conical, and interlocking, similar to those of nothosaurs and plesiosaurs, and well-adapted to catching moving prey. Further evidence is that hooklets from cephalopod tentacles and fish scales have been found in the belly regions of some skeletons.
 |
| a lone Doswellia stalks the Carnian Triassic landscape |
Another archosaurimorph (this one terrestrial) was
Trilophosaurus from North America. This herbivore reached up to eight feet long and was short with an unusually heavy skull equipped with massive, broad, flattened cheek teeth with shearing surfaces for cutting tough plant material.
Doswellia was a low and heavily-built carnivore with a wide, flattened head and narrow jaws. It had a box-like rib-cage surrounded by rows of bony plates (or osteoderms). It had pointed teeth, a long snout, and upward-pointing eyes, all characteristics that hint towards it being an aquatic carnivore (though these features are also seen in
Parasuchus, a phytosaur that preyed on terrestrial reptiles).
Doswellia likely enjoyed bivalves, crustaceans, and insects. Though front limbs haven't been discovered, it's theorized that it may have been able to partially burrow so that its soft parts were underground with just its armor exposed, a tactic used by modern armadillos. Another archosauriform,
Hyperodapedon, was heavily-built, stocky, and reached just over four feet in length. It's been classified as an early member of the rhynchosaurs (stocky, beaked, herbivorous reptiles). Apart from its beak, it had several rows of heavy teeth on each side of the upper jaw and a single row on each side of the lower jaw; together, these created a powerful chopping action when it ate. It's believed to have been herbivorous, subsisting primarily on seed ferns; when seed ferns withered away at the end of the Triassic,
Hyperodapedon withered away with them.
 |
| an artist's rendition of Hyperodapedon |
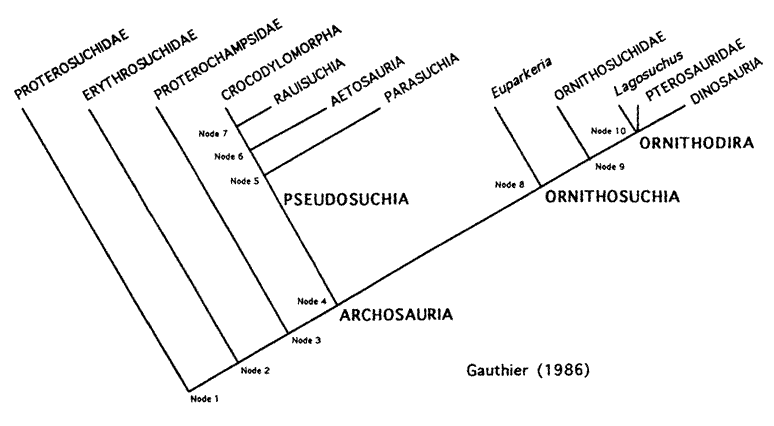
The Carnian saw the radiation of the pseudosuchians (or 'false crocodiles'). The group Archosauria splits into two sub-groups: the pseudosuchians and the ornithosuchians. The pseudosuchians include all archosaurs more closely related to modern crocodiles than to birds; the ornithosuchians, which split after the pseudosuchians, includes all archosaurs more closely related to birds than crocodiles. Pseudosuchia, of interest here, further dovetails into an assemblage of different types of creatures: the armored and herbivorous aetosaurs, the aquatic (or, at least, semi-aquatic) phytosaurs, the ornithosuchians (who resemble later ornithosuchids but who are nonetheless more closely related to crocodiles), the erythrosuchids, the terrifying rauisuchians, and the crocodylomorphs. The latter group were the only pseudosuchids to survive the Triassic-Jurassic extinction event, and their descendants are among us today as alligators, crocodiles, caimans, and gharials. Below is a cladogram detailing the 'pseudosuchid family tree':

 |
| Desmatosuchus |
Desmatosuchus was a heavily-armored aetosaur. Aetosaurs were heavily-armored, medium to large-sized Late Triassic herbivorous archosaurs. They had small heads, upturned snouts, erect limbs, and bodies that sported plate-like scutes. The two distinct subdivisions - Desmatosuchinae and Aetosaurinae - are based on differences in the bony scutes of the two groups. More than twenty different species have been named.
Desmatosuchus' carapace was made up of two rows of median scutes surrounded by two more rows of lateral scutes. The front spine was much larger than the rest, around twenty-eight centimeters, and recurved.
Desmatosuchus was the only aetosaur known to possess such spines.
 |
| Rutiodon amid several smaller Hesperosuchus |
The aquatic phytosaurs radiated during the Carnian.
Plaeorhinus grew up to eight feet long, but
Rutiodon grew even larger, between nine and twenty-five feet in length.
Rutiodon looked a lot like a crocodile, but its nostrils were positioned far back on the head, close to the eyes, instead of on the tip of the snout. It had enlarged front teeth and a narrow jaw resembling modern gharials. It was a carnivore that fed mostly on fish, but it likely snagged land animals that dared come down to the waterfront to drink. It had armored plates on its back, flanks, and tail.
 |
| Ornithosuchus |
Ornithosuchus ('bird crocodile') resembled the ornithosuchians but was more closely related to crocodiles. It was able to walk on its hind legs, though it probably spent most of its time on all fours. Its skull resembled those of theropod dinosaurs, but it had primitive features such as the presence of five toes on each foot and a double row of armored plates along its back. It could reach up to thirteen feet in length and was likely a scavenger.
 |
| Saurosuchus in a Carnian tropical forest |
Saurosuchus ('lizard crocodile') was a predecessor to the crocodilians that grew up to twenty-three feet long. It walked on four fully-erect limbs. It had a deep skull with large, recurved, serrated teeth. The skull was wide in the back and narrowed in front of the eyes. Its skull roof and maxilla were pitted; this pitting is also seen in aquatic phytosaurs and crocodilians, but the ridges and grooves in these animals were more extensive. The frontal bones, at the top of the skull, were enlarged to form thick ridges over the eyes.
Saurosuchus had a strong neck and two rows of dorsal osteoderms running along its back. A closer relative to modern crocodiles was
Hesperosuchus, which is considered a crocodylomorph (part of the family line that would eventually lead to modern crocodiles and their attendants). Ironically, in outward appearance it looked less like a crocodile than other archosaurs that didn't lead to modern crocodiles. It was a small terrestrial critter that could run fast from predators and against prey. It's been found in areas that, in the Carnian, were surrounded by lakes and streams. Though terrestrial, it kept close to water and likely fed on freshwater fish.
 |
| Postosuchus chomping down on an early dinosaur |
Postosuchus is one of the most famous rauisuchians (predatory reptiles that grew up to twenty feet or more in length and which were the apex terrestrial predators of the Upper Triassic).
Postosuchus reached between thirteen and sixteen feet, was heavily-built and robust, and had a deep skull and long tail. Many paleontologists believe
Postosuchus was bipedal given the shortness of the forelimbs compared to the hind-limbs, the very small hands, and the measurement of the vertebrae.
Postosuchus was one of the top tier carnivores of its day, literally a head and shoulders above the rest. Adults reached up to four feet in height and weighed around a quarter of a ton. Its massively-built skull was filled with dagger-like teeth, and it had osteoderms on its back, neck, and possibly above (or under) the tail.
Postosuchus lived in the tropical environment in Pangaea’s exterior; this moist, warm region consisted of ferns, gymnosperms, and cicads. It prowled a region filled with lakes and rivers and was particularly common in the Upper Triassic uplands along with
Coelophysis and
Desmatosuchus. As a top predator it would’ve fed on super-salamanders, other archosaurs, and early dinosaurs.
 |
| an artist's rendition of a furry Carniadactylus |
The first pterosaur,
Carniadactylus, appeared during the Carnian. It was closely related to the better-known Norian-age
Eudimorphodon, though it was much smaller. Like
Eudimorphodon, it had complex multi-cusped teeth. The significant size difference has been attributed to them occupying different niches and relying on different food surfaces. This is supported by studies of their teeth:
Carniadactylus’ teeth show little to no wear, while the fish-eating
Eudimorphodon has worn-down teeth from chewing food. This has led scientists to speculate that the smaller
Carniadactylus probably fed on smaller, soft-bodied prey like worms and insect larvae.
The Carnian of the Upper Triassic is where we find the first (agreed upon) true dinosaur,
Eoraptor, along with a slew of emergent dinosaurs. Dinosaurs are distinct from other archosaurs based upon a number of anatomical features found in their skeletons, not least of all their stance: all dinosaurs had an erect, pillar-like stance with their legs positioned directly beneath their bodies. All dinosaurs were terrestrial in that none were aquatic (though some spent time in water, such as the much later
Suchomimus and
Spinosaurus), and they are distinguished from Mesozoic marine reptiles (such as the ichthyosaurs, plesiosaurs, and mosasaurs) and from the Mesozoic flying reptiles (the pterosaurs and their kin). Though it was believed for a long time that no dinosaurs flew, by the end of the Mesozoic (if not as early as the middle Jurassic or even, as some argue, the Late Triassic) some had taken to the skies as birds. The earliest dinosaurs, such as
Eoraptor, were bidepal carnivores (‘theropods’), but early on a divergent branch of dinosaurs became herbivorous. Most of the Upper Triassic carnivores and herbivores were ‘saurischians,’ in that they had lizard-like hips; the other group of dinosaurs, ornithischians, had bird-like hips.
Pisanosaurus, of the Carnian, is considered by many to be the first true ornithischian dinosaur. Carnivorous theropods would remain small during the Triassic, but herbivorous ‘prosauropod’ dinosaurs (the ‘forerunners’ of the titanic Jurassic long-necked sauropods) would become the biggest Triassic herbivores by the end of the period. Though Carnian dinosaurs radiated, they were nonetheless outperformed and outmatched by their archosaur cousins and were in no way the dominant Triassic terrestrial lifeforms. Some Carnian dinosaurs include
Caseosaurus,
Chindesaurus,
Euskelosaurus,
Herrerasaurus,
Pisanosaurus,
Eoraptor, and
Staurikosaurus. We will examine a few of them.
 |
| Eoraptor |
Eoraptor lived in what is modern-day Argentina. It had a slender body that reached about three feet in length and weighed around twenty pounds. Its skull was lightly built and similar to those of the later coelophysoids. Like all dinosaurs it had a digitigrade stance, walking upright on its toes, and it stood upright on its hind legs. All of its long bones had hollow shafts and hollow central vertebrae. It had five digits on each ‘hand,’ the three longest of which ended in large claws presumably used to handle prey. It had heterodont teeth (meaning it had multiple types of teeth), so it’s speculated that it was omnivorous. It had serrated, recurved teeth in its upper jaws (like those of later theropods) and leaf-shaped teeth in the lower jaw (like those of prosauropods). It was a fast runner and could use its claws and teeth to tear prey apart. Unlike later theropods, it lacked a sliding joint at the articulation of the lower jaw that would enable it to hold large prey. Its remains have been found in what was once a volcanically-active floodplain covered by forests with a warm and humid climate subject to monsoons. It lived among ferns, horsetails, and giant conifers of the highland forests that stretched along the banks of rivers. It was likely preyed upon by another (and much larger) early theropod,
Herrerasaurus.
Eoraptor would’ve shared its environment with Carnian therapsids, rauisuchians, and archosaurs. Dinosaurs, of which
Eoraptor was a part, represent only six percent of the fauna found in Argentina. It certainly wasn’t the dominant creature.
 |
| Herrerasaurus |
Herrerasaurus far outweighed
Eoraptor at twenty feet in length and up to 750 pounds in weight. This Carnian predator was lightly built with a long tail and a relatively small head. It was fully bipedal with strong hind limbs and long feet. The feet had five toes, but only the middle three bore weight. The tail, partially stiffened by overlapping vertebral projections, balanced the body and was an adaptation for speed.
Herrerasaurus’ upper arms and forearms were rather short, but the hands were elongated. The first two fingers and the thumb ended in curved, sharp claws used for slashing and gripping prey. The other fingers were small stubs without claws. Its long, narrow skull lacked most of the specializations seen in later dinosaurs, but it had a flexible joint in the lower jaw that could slide back and forth to deliver a grasping bite. Its jaws had large serrated teeth for biting and tearing, and its slender neck was flexible. It lived in the same environment as
Eoraptor and likely preyed upon super-salamanders and herbivorous dicynodonts; it would’ve competed with rauisuchians and phytosaurs for food.
 |
| Staurikosaurus |
Staurikosaurus was a small and agile predator that reached about seven and a half feet in length, about two and a half feet in height, and clocked in around sixty pounds.
Staurikosaurus is based on fragmentary material, but enough has been found to know that it was a quick-running carnivore with serrated teeth that curved back towards the throat, enabling it to catch and hold onto prey; though this teeth feature would disappear in later theropods, it was common among those of the Upper Triassic. A sliding joint in its jaw allowed the jaw to move backwards and forwards, as well as up and down, so that smaller prey could be worked backwards towards the throat. It had a long tail that it held off the ground as it ran; the rear part of its tail was stiffened and acted as a dynamic stabilizer enabling the creature to run and leap with precise agility. It would’ve preyed on small and medium-sized animals.
 |
| the early prosauropod Euskelosaurus |
Euskelosaurus was an herbivorous prosauropod that appeared in the Carnian and survived into the early Jurassic.
Euskelosaurus was the first dinosaur to be discovered in Africa, and the number of specimens found in South African sandstones imply it had been a staple of the area’s ecosystem.
Euskelosaurus foreshadowed the giant sauropods of the Jurassic, with adults reaching up to forty feet long head-to-tail (that estimate may change depending on the head; though scientists have a good idea of what it looked like, a skull has yet to be found). Its name means “good-limbed lizard,” and it strikes a chord of irony: the shaft of its thigh bone was twisted, which may have rendered it bow-legged! Because dinosaurs positioned their legs underneath their bodies, this anatomical feature is a bit of a mystery. Some have proposed that
Euskelosaurus’s earliest versions had an erect posture, but as its body grew larger, vastly exceeding the size of its proto-dinosaurian ancestors, its femur twisted in an evolutionary attempt to handle the added weight. This evolutionary sideshow would’ve been inefficient compared to keeping an erect stance, and perhaps this is why
Euskelosaurus doesn’t seem to have made it past the Lower Jurassic.
 |
| two Pisanosaurus flee a hungry rauisuchian |
Pisanosaurus was a small and lightly-built herbivore that grew to about three feet long. It lived in the same environment as
Eoraptor and
Herrerasaurus and likely spent a lot of time running from them and other predators.
Pisanosaurus’ place on the dinosaur family tree was hotly debated for nearly four decades, but in 2017, after meticulous study and argumentation, most paleontologists agreed that it’s the earliest-known ornithischian dinosaur. Most Triassic dinosaurs were saurischian (lizard-hipped) and would radiate into the theropods and prosauropods (and, later into the Jurassic, sauropods); the ornithischian dinosaurs, despite their austere beginnings with
Pisanosaurus, would blossom throughout the later Mesozoic into stegosaurs, ankylosaurs, hadrosaurs and lambeosaurs, ceratopsians and pachycephalosaurs. Though most paleontologists embrace
Pisanosaurus as an ornithischian, others believe it was actually a silesaurid, a type of archosaur that is the closest known relative to the first dinosaurs.
~ The Norian Stage ~
237 to 208.5 mya
Upper Triassic
The base of the Norian begins at the ammonite biozones of
Klamathites macrolobatus and
Stikinoceras kerri; the top of the Norian is the first appearance of the ammonite species
Cochloceras amoenum. The Norian stage of the Upper Triassic witnessed the rise of the herbivorous sauropodomorph dinosaurs and the extinction of several large herbivorous dicynodonts. It’s unknown if the rise of sauropodomorph dinosaurs is what led to the extinction of the biggest dicynodonts (perhaps the dicynodonts couldn’t compete?) or if the dicynodonts suffered for other reasons and the sauropodomorphs rushed to fill the niches they left vacant. Some argue that the dicynodont decline was due to a rise in aridity that pushed the moist environments dicynodonts favored even more to the Pangaean fringes; in this case the dicynodonts couldn’t cope with environmental changes, and the sauropodomorphs were more able to adapt and shouldered their way towards dominance. Nevertheless, dinosaurs remained a ‘fringe group’ in the Upper Triassic, reaching at most twenty percent of earth’s terrestrial life forms.
The Norian saw continued success with several types of life-forms, but oddly enough many types of creatures remained evolutionary stagnant while dinosaurs and their kin radiated. This has not been adequately explained, but the most likely explanation is that the life-forms that the dominant non-dinosaurian life forms that evolved during the Carnian didn’t need to evolve to keep up the pace in the Norian. Absent environmental factors, life could continue relatively as normal. The carnivorous dinosaurs, however, were constantly pushed to outperform their non-dinosaur counterparts; this would lead to swift radiation among the theropods. The herbivorous dinosaurs, particularly the prosauropods, got a boost in evolution when the niches formerly dominated by dicynodonts were left vacant. Thus while there are few emergent forms of many non-dinosaur archosaurs, the dinosaurs saw a panoply of swift evolution during the Norian. The Norian also saw continued dominance of the ichthyosaurs at sea, a radiation of pterosaurs, and the rise of turtles.
 |
| Himalayasaurus, an ichthyosaur cousin of the gigantic Shonisaurus |
In the Norian seas,
Shonisaurus continued to dominate with very little change. In the modern Himalayas, another giant ichthyosaur,
Himalayasaurus, flourished. The Himalayan Mountains are a recent addition to our planet’s geography; in the late Triassic, the Himalayas were a smooth, shallow sea.
Himalayasaurus resembled its cousin
Shonisaurus, but its teeth were distinguished by the cutting-edges on its flattened tooth crowns (a feature unique to this ichthyosaur). Its teeth suggest that it roamed the ancient shorelines feeding on terrestrial or semi-aquatic prey; in this way it would’ve acted like modern killer whales. Another development in the seas was the emergence of the placodont
Psephoderma with its triangular skull, pointed rostra, and extensive carapace.
 |
| Eudimorphodon |
Pterosaurs radiated in the Norian skies. The most famous of all Norian pterosaurs was
Eudimorphodon – not only because of the wealth of specimens (including numerous juveniles) but also because of an array of teeth that would’ve been an orthodontist’s worst nightmare. It resembled the Carnian’s
Carniadactylus but was larger with a wingspan of just over three feet. It weighed around twenty pounds, and its gargantuan fourth finger attached to the membrane making up its wing. Its panoply of heterodont teeth – packed to the gills with 110 in a jaw only six centimeters long! – make it notable among pterosaurs; most pterosaurs either had simple teeth or lacked them altogether. The front of its jaw had fangs, four per side in the upper jaw and two per side in the lower jaw, and these fangs gave way to a line of twenty-five multi-pointed teeth in the upper jaw and twenty-six in the lower jaw. This array of teeth suggests a piscivorous (or fish) diet, and this assumption was verified by the discovery of bony fish remains in the stomach regions of one fossilized
Eudimorphodon. It likely also fed on insects and hard-shelled invertebrates. The end of its bony tail may have been diamond-shaped like the laer
Rhamphorhynchus; if so, it may have helped
Eudimorphodon steer while in the air.
 |
| Preondactylus |
A slightly-smaller pterosaur of the Norian was
Peteinosaurus. It had a long tail stiffened by vertebral extensions, a feature common among Triassic pterosaurs. Its bones were strong but lightly-built, and it had three types of conical teeth. It likely fed on insects. Another Norian pterosaur was
Preondactylus. It was larger than both
Eudimorphodon and
Peteinosaurus with a wingspan of up to one and a half meters, and like both it was found in Italy. It had a long tail and relatively long legs. It had single cusp teeth (meaning they had one point on each tooth) and likely fed on fish and insects. One specimen had a sad turn of luck when it dived for a fish and was instead eaten by one: this specimen was discovered in the gastric pellet of a predatory fish that had vomited up the pterosaur’s indigestible parts (and which were later fossilized). The artist’s rendition of
Preondactylus to the left shows it covered in pycnofeathers (a type of early feather); though we tend to imagine pterosaurs as leathery-winged creatures, the discovery of feathered pterosaurs in China has contested this notion. The fossil beds in China are of a sort that enabled many exquisite features to fossilize, and we know that at least some pterosaurs had feathers. We also know that many dinosaurs, of both ornithischian and saurischian stock, had feathers; because dinosaurs and pterosaurs diverged early on in archosaur history, it’s surmised by many that early feathers originated in basal archosaurs and passed on to pterosaurs, dinosaurs, and (in a highly-specialized form) modern birds. While the recent finds don’t tell us that all pterosaurs had feathers, we can at least surmise that all had the genetic potential for some time of feathered augmentation. These early feathers (unlike those found in modern birds) weren’t designed for flight; pterosaur flight, while working on the same mechanics as birds, was engineered with membrane wings rather than feathered wings. In their case, the feathers would’ve likely been for show – a way to warn rivals or attract mates.
 |
| a pair of mating Proganochelys |
It was during the Norian that a modern animal came upon the seen: the first of the family Chelonia, better-known as turtles. They appeared like a lightning bolt from space and dominated much of the fauna in the blink of an eye. Two species from the Norian are known:
Proganochelys quendstedti and
Proterochersis robusta. The first was a large turtle that reached about three feet in length. It had a solid, robust skull with a toothless jaw and horny beak (though, as a primitive form, it retained tiny teeth on the roof of its mouth). Its broad domed shell had small spikes that projected outwards, and the shell’s openings were too small to permit the head and legs to be drawn in. The exposed limbs were protected by the spiked plates, and the neck had its own assemblage of spiny scales.
Proterochersis robusta is more advanced than
Proganochelys, despite appearing earlier than its compatriot.
Proterochersis had larger openings in its shell that allowed the limbs to retract (though turtles wouldn’t develop the ability to retract their heads until the Cretaceous; and semi- and fully-aquatic turtles wouldn’t show up until the Jurassic). The sudden appearance of these two very different species implies a ‘ghost-lineage’; in other words, it’s speculated that there were ‘turtle forebears’ that existed, perhaps, in the Carnian. Maybe the disappearance of rivals or predators in the early Norian enabled these ‘underground’ species to radiate.
 |
| Teratosaurus, a Norian rauisuchian |
Now we will examine developments among the Norian archosaurs before turning our attention to the emerging dinosaurs. The heavily-armored and herbivorous
Aetosaurus – considered to be one of the most primitive of the aetosaurs – evolved into two species and dominated northwest to middle-north Pangaea. Their armor would’ve been helpful against predatory archosaurs such as the twenty-foot-long rauisuchian
Teratosaurus. As far as crocodylomorphs go,
Saltoposuchus radiated during the Norian in what is now Europe. This close relative to modern crocodiles didn’t look much like a crocodile at all; it only reached about three to five feet and has been described as ‘a greyhound dog with scales and a long tail.’ It had fully erect slender limbs, a slender body, and long legs and a long, pointed skull with pointed teeth. It was a terrestrial creature designed for running at fast speeds, and many scientists speculate that it hunted in packs. It likely dined on small lizards, insects, and early mammals (such as
Eozostrodon,
Kuehneotherium,
Morganucodon, and
Sinocodon).
 |
| the agile relative of modern crocs, Saltoposuchus |
 |
| Plateosaurus, one of the largest dinosaurs of the Norian |
Archosaurs didn’t see much radiation during the Norian – except for the dinosaurs. Dinosaurs exploded in diversity, reaching epic proportions and becoming quite specialized. This doesn’t mean that they were particularly numerous – as already mentioned, they remained a minor subset of the ecosystems despite their variation. The largest of them all was
Plateosaurus, which lived in Late Triassic Europe and could reach up to 30 feet in length. This dinosaur seems to have had a penchant for fossilization, as paleontologists have studied over a hundred skeletons found in the Triassic sandstones of Germany, France, and Switzerland. It had a pear-shaped body made longer by its long neck and tail, with the tail making up half its total length. Its jaws were filled with small, coarsely-serrated, and leaf-shaped teeth; the low-slung hinge of its lower jaw gave its jaw muscles greater leverage to produce a powerful bite. Small ridges of bone around its mouth supported fleshy cheek pouches in life: the front teeth stripped the leaves, and the pouch kept vegetation from falling out of the mouth before it could chew and/or swallow, thus preventing needless waste (an animal of
Plateosaurus’ size needed all the food it could get!). Its front legs were shorter than its hind-legs, and it may have been able to rear up on its hind legs not only to reach higher foliage but to run from predators: because of the length of the lower leg bones, some scientists speculate that it could have reached top speeds running on two legs. The hind limbs had slightly flexed knees and ankles, and its feet were digitigrade, meaning they walked on their toes like modern birds. Its eyes were directed to the sides, rather than to the front (like predators), providing all-round vision to keep a wary eye out for threats. Some
Plateosaurus skulls have preserved sclerotic rings (rings of bony plates that protected the eyes); by comparing the scleral rings and orbit sizes of
Plateosaurus and modern birds and reptiles, scientists have suggested that
Plateosaurus was cathemeral, meaning that it was active at any time during the day or night, depending on circumstances. Recent studies estimate that Plateosaurs tended to live between 12-20 years, but their maximum age is unknown (though one specimen seems to have been around 27 years of age). Fossil beds imply that
Plateosaurus was a social animal that likely cared for its young. One can imagine massive herds of
Plateosaurus roaming the Upper Triassic plains like the American buffalo of the 1800s.
 |
| Thecodontosaurus, a small prosauropod |
Another notable herbivorous dinosaur was the prosauropod
Thecodontosaurus, which appeared in the Norian and became widespread in the following Rhaetian. It was bipedal and averaged about four feet in length (though one particular individual reached up to eight feet long), one foot in height, and weighed about twenty-four pounds. Most of its skeleton can be reconstructed, except for the front part of its skull. From the fossils available, we know that
Thecodontosaurus had a rather short neck and a large skull with large eyes. Its jaws contained many small- to medium-sized serrated, leaf-shaped teeth. Its hands and feet each had five digits, and the hands were long and narrow with an extended claw on each. Its front limbs were much smaller than its hind limbs, and the tail comprised more than half the length of its body. Continuing research on
Thecodontosaurus has shown that there were two main morphologies, one much more robust than the other. Some paleontologists believe these two morphologies represent separate species; others argue that it’s a matter of sexual dimorphism, which would imply that
Thecodontosaurus engaged in gregarious (or social) behaviors like the much-larger prosauropod
Plateosaurus.
 |
| a lone Coelophysis on the prowl for food |
Theropods (carnivorous bipedal dinosaurs) made leaps and bounds during the Norian.
Coelophysis has become archetypical for Triassic carnivorous dinosaurs, thanks to the wealth of specimens found at Ghost Ranch, New Mexico.
Coelophysis grew up to about ten feet in length and was a fast, agile runner. It may have traveled in packs, coordinating to bring down larger prey, but most of its diet likely consisted of creatures smaller than itself. It used to be believed that
Coelophysis was cannibalistic because of supposed remains of juveniles found in the stomach regions of adults, but further analysis revealed these remains belonged to a crocodylomorph.
Coelophysis is more advanced than its Carnian forebears
Herrerasaurus and
Eoraptor, and it's the first theropod to have a wishbone (which would become an important anatomical structure for later birds). A larger theropod that likely fed on larger prey was
Liliensternus.
Liliensternus could grow up to nearly seventeen feet in length and, like
Coelophysis, had a penchant for fossilization. It’s one of the better-known Triassic theropods from Europe – and one of the largest Triassic theropods. It was an active bipedal carnivore that likely preyed upon the larger herbivores in its environment, including archosaurian aetosaurs and the prosauropod
Plateosaurus. It lived in ancient floodplains that were abundant with reptiles, therapsids, and a favorite roaming ground of
Plateosaurus. Its slashing teeth would’ve enabled it to take down large prey, and its swift speed would enable it to chase down swift creatures.
 |
| a pair of Liliensternus face-off with a Plateosaurus |
~ The Rhaetian Stage ~
208.5 to 201.3 mya
Upper Triassic
The bottom of the Rhaetian is the ammonite biozones of
Sagenites reticulates and
Cochloceras amoenum; the top of the Rhaetian, and the beginning of the Jurassic Period, is the first appearance of the ammonite genus
Psiloceras. On land, ecosystems continued operating with little change, except that in most of Pangaea sauropodomorphs dominated at the expense of the herbivorous aetosaurs. The armored archosaurian aetosaurs were pushed to the northwestern fringes of Pangaea where they lived among phytosaurs; throughout the rest of Pangaea, the prosauropods and first sauropods had the herbivorous niches on lockdown, and the carnivorous theropod dinosaurs flourished.
 |
| a pair of Leptonectes |
The biggest changes in this stage happened in the oceans: the nothosaurs, which emerged earlier on in the Triassic, were driven to extinction by the emergent plesiosaurs; and the whale-like ichthyosaurs – such as
Shonisaurus,
Shastasaurus, and
Himalayasaurus – were replaced by their dolphin-like cousins, such as
Leptonectes. The first plesiosaurs were smaller than the Loch Ness monsters most of us conjure in our minds. One of the earliest-known plesiosaurs was
Thalassiodracon, which reached about six to nine feet in length; another was
Eretmosaurus, which reached about nine feet long. Though these early plesiosaurs had long necks, they’re classified as ‘pliosaurs’. Plesiosaurs have been divided into two camps: the long-necked plesiosaurs proper and the short-necked pliosaurs. Despite having long necks, the earliest plesiosaurs shared more dominant morphologies with pliosaurs, and thus they’re classed in the ‘short-necked’ category. Plesiosaurs would diversify and grow larger throughout the rest of the Mesozoic before going extinct at the end of the Cretaceous.
 |
| an early plesiosaur |
 |
| the 'super-croc' Redondosaurus takes down a sauropodomorph |
As already mentioned, predatory phytosaurs and herbivorous aetosaurs remained dominant; elsewhere, sauropodomorph dinosaurs – such as the prosauropods – dominated. Theropods, despite austere beginnings earlier in the Upper Triassic, bloomed like wildflowers. Prosauropods dominated northern Pangaea, and the first basal sauropods emerged in the south (though the southern prosauropod
Riojasaurus remained dominant in the southern niches). Large herbivores tended to stay in the valleys and floodplains, while smaller prosauropods – such as
Thecodontosaurus and its kin – carved out the uplands for themselves. The Rhaetian was the twilight of the archosaurs (with the exceptions of dinosaurs and crocodylomorphs), but they went out with a bang.
Redondosaurus was a large, advanced phytosaur; and the fast-moving, fully-erect
Riojasuchus was a medium-sized terrestrial carnivore about six feet in length. Overwhelming both of these archosaurs was one of the last (and largest) rauisuchians:
Fasolasuchus’ skull measured three feet long, and it likely grew close to thirty feet in length. Scientists speculate that
Fasolasuchus evolved to specialize on taking down large sauropodomorphs.
 |
| the rauisuchian Fasolasuchus threatens a sauropodomorph |
 |
| a floundering Riojasaurus is threatened by a rauisuchian |
One of the largest prosauropods,
Riojasaurus, was the ‘plateosaur of the south.’
Riojasaurus was larger than its northern cousin, stretching up to thirty-six feet in length, and was likely fully quadrupedal. Its backbone was punctuated by the presence of hollow, air-filled cavities that kept the spine from being too heavy; these hollow spaces are rare in prosauropods but predominant in sauropods. The fact that
Riojasaurus had these hollow cavities has prompted many scientists to claim it as a ‘second cousin’ to the sauropods. It isn’t surprising, then, that the first basal sauropods emerged alongside
Riojasaurus in southern Pangaea.
Isanosaurus was the first pre-Jurassic sauropod discovered, but it was outdone by
Antetonitrus. Cladistic studies on these early sauropods indicate that sauropods emerged much earlier than these specimens appear. Another Rhaetian sauropod was the twenty-seven-foot long
Camelotia. Though the early sauropods didn’t outsize their larger prosauropod cousins, this would change in the Jurassic as sauropods blossomed into titanic, awe-inspiring sizes.
 |
| a herd of Isanosaurus, one of the first basal sauropods |
Though theropods radiated and flourished throughout the Rhaetian, they didn’t become much larger than they did in the Norian (that story would change in the Jurassic, however).
Agrosaurus reached ten feet in length, and
Zupaysaurus grew to about thirteen feet long. At one time
Zupaysaurus was considered an early tetanuran (the type of theropod to which dromaeosaurs, tyrannosaurs, and birds belong), but further studies showed it to be a coelophysoid with some hit-or-miss tetanuran characteristics (a matter of convergent evolution). It likely fed on smaller sauropodomorphs (or even on juvenile sauropods), and it had a long neck with a head decorated by paired crests likely for mating theatrics.
 |
| Zupaysaurus |
The Rhaetian was the last stage of the Triassic, and it ended with a bang (or, rather, three of them). The Permian Period saw the formation of the supercontinent Pangaea, and the Late Triassic saw the beginning of its breakup. As Pangaea began to split (the upper portion would become ‘Laurasia’ and the southern portion ‘Gondwana’), it stretched at the seams, creating ‘rift basins.’ The shallow bowls of stretched earth were often filled with water, creating lakes and rivers; many would seasonally dry out, resulting in fantastic ‘bone beds’ for future paleontologists. As the stretching intensified, the ripping began. Pangaea ruptured along the modern-day U.S. eastern seaboard (the Palisades in New York are remnants of this split), and the rift basins were inundated by volcanic lava flows. There would be four cycles of mass eruptions before Pangaea finally split down the middle, and the process took about half a million years. These volcanic ‘pulses’ weren’t meager little things: they sent tsunamis of lava surging across the Late Triassic landscape. This is no exaggeration, as some of the lava flows reached up to three thousand feet in thickness. These lava tsunamis could’ve buried the Empire State Building twice over.
Three million square miles of central Pangaea were buried by lava, and the effect on climate was catastrophic. Survival wasn’t just a matter of hoofing it away from the split; the split itself initiated a domino-effect of rising greenhouse gases that makes today’s climate change scare look like amateur hour. The climate change would wipe out thirty percent of earth’s species and help the emergent dinosaurs break out of their slump and diversify into the dominant creatures of the Mesozoic. The intense volcanism spewed carbon dioxide, a ‘greenhouse gas’ (so-called because it warms the planet), into the atmosphere. As the oceans warmed due to the increased carbon dioxide, clathrates melted; clathrates are chunks of ice buried in the seafloor in which methane is trapped, and when clathrates melt, the trapped methane – another greenhouse gas – is released. The unchained methane piggybacked on the carbon dioxide, and the greenhouse gases in the atmosphere tripled within a few tens of thousands of years. The planet’s temperature warmed at a nauseating pace, and life struggled to keep tempo. The ocean became more acidic, resulting in the collapse of marine food webs; shellfish were particularly hard-hit. The climate became more variable: periods of intense heat were followed by rapid cooling, megamonsoons became far more severe, and coastal regions became wetter while interior regions became drier. The hotter temperatures led to the death of ninety-five percent of plant life; herbivores suffered from the lack of food, and the carnivores that preyed upon them suffered in turn. Most reptiles, amphibians, and early mammals died out.
The supersalamander amphibians? They were gone.
The piglike dicynodonts? Gone.
The beaked rhynchosaurs? Gone.
The long-snouted phytosaurs, the tank-like aetosaurs, and the apex-predator rauisuchians were gone, gone, GONE.
And the pseudosuchians? They were almost wiped out. The only traces of them were a few types of primitive creatures that would eventually evolve into modern alligators and crocodiles, but they would never again enjoy the success they’d claimed in the Triassic.










































No comments:
Post a Comment